![]()
Caroline Marques Castro1, José Francisco Montenegro Valls2
& Cláudio Takao Karia3
1Pesquisador da Embrapa Clima Temperado,
BR 392, Km 078, C. P. 403, Pelotas - RS, Brasil. CEP. 96001-970.
(
caroline@cpact.embrapa.br
)
2Pesquisador da Embrapa Recursos Genéticos e Biotecnologia,
Av. W3 Norte (final) s/n, Ed. Sede Embrapa, CEP. 02372, Brasília – DF, Brasil. CEP. 70770-900. (
valls@cenargen.embrapa.br
)
3Pesquisador da Embrapa Cerrados,
BR 020, Km 018, C. P. 403, Planaltina - DF, Brasil. CEP. 73301-970.
(
karia@cpac.embrapa.br
)
El presente artículo, es el segundo de una serie, que Caroline Marques Castro presentó al Instituto de Biociencias, Campus de Botucatu, de la Universidad Estadual de São Paulo UNESP, como parte de los requisitos necesarios para la obtención del título de Doctor en Ciencias Biológicas - Genética.
RESUMO
 O interesse em Arachis pintoi vem crescendo, com mais de dez cultivares lançadas nos últimos 12 anos, em vários países. Grande variabilidade é encontrada no germoplasma desta espécie com relação aos componentes biológicos da produção de forragem e de sementes. O presente trabalho teve como objetivo identificar, em três populações F2 de cruzamentos intraespecíficos de Arachis pintoi, as correlações entre os caracteres comprimento e largura dos folíolos basal e distal, percentagem de cobertura do solo, comprimento do entrenó, espessura do estolho, número de estolhos, produção de matéria seca, número de flores/m2, número de sementes/m2, peso médio de sementes e comprimento e densidade de pêlos no ápice dos estigmas das flores. As duas avaliações de produção de matéria seca apresentaram correlações positivas com todos os caracteres vegetativos avaliados. Os altos valores de produção de matéria seca, percentagem de cobertura do solo e de espessura do estolho, foram compatíveis com os maiores valores de número de sementes/m2. Entretanto, foi negativa a correlação entre o número de sementes/m2 e o comprimento de pêlos no ápice do estigma, enquanto que com a densidade de pêlos na superfície estigmática foi positiva. Os dados mostraram a possibilidade de agregar em um único genótipo, características desejáveis de cobertura do solo, produção de forragem e de sementes. No entanto, é importante considerar o comprimento de pêlos no ápice do estigma como uma barreira no processo de polinização.
O interesse em Arachis pintoi vem crescendo, com mais de dez cultivares lançadas nos últimos 12 anos, em vários países. Grande variabilidade é encontrada no germoplasma desta espécie com relação aos componentes biológicos da produção de forragem e de sementes. O presente trabalho teve como objetivo identificar, em três populações F2 de cruzamentos intraespecíficos de Arachis pintoi, as correlações entre os caracteres comprimento e largura dos folíolos basal e distal, percentagem de cobertura do solo, comprimento do entrenó, espessura do estolho, número de estolhos, produção de matéria seca, número de flores/m2, número de sementes/m2, peso médio de sementes e comprimento e densidade de pêlos no ápice dos estigmas das flores. As duas avaliações de produção de matéria seca apresentaram correlações positivas com todos os caracteres vegetativos avaliados. Os altos valores de produção de matéria seca, percentagem de cobertura do solo e de espessura do estolho, foram compatíveis com os maiores valores de número de sementes/m2. Entretanto, foi negativa a correlação entre o número de sementes/m2 e o comprimento de pêlos no ápice do estigma, enquanto que com a densidade de pêlos na superfície estigmática foi positiva. Os dados mostraram a possibilidade de agregar em um único genótipo, características desejáveis de cobertura do solo, produção de forragem e de sementes. No entanto, é importante considerar o comprimento de pêlos no ápice do estigma como uma barreira no processo de polinização.
Abstract
The interest in Arachis pintoi has grown, and more than 10 cultivars were released in various countries in the last 12 years. Germplasm of this species presents great variability with regard to biological components of seed and forage production. The present work aimed to identify the correlations among these characters, in three F2 progenies of intraspecific crosses of Arachis pintoi. Length and width of basal and distal leaflets, percentage of soil mulch, length of internodes, thickness and number of stolons, dry matter, number of flowers and seeds/m2, average weight of seeds and length and density of pilosity in the top of flower stigmas were evaluated. The two evaluations of dry matter production presented positive correlations with all other vegetative characters studied. The high values of dry matter production, percentages of soil mulch and stolon thickness were compatible with the highest values of seed number/m2. However, this variable and length of hair in the top of the stigma showed negative correlation, while it was positive when compared to density of hair in stigmatic surface. The data showed that is possible aggregate desirable characteristics of soil mulch, seed and forage production in just one genotype. Nevertheless, it is important to consider the length of hair in the top of stigma as a barrier in the pollination process.
Palavras-chave: amendoim forrageiro, melhoramento de plantas
Key words: forage peanut, plant breeding
Introdução
 A plasticidade no uso de Arachis pintoi como planta forrageira para pastejo direto, para a produção de feno e silagem, para uso paisagístico e em cultivos de cobertura faz desta espécie uma leguminosa ideal para os trópicos latino-americano e mundial (PIZARRO, 2001b). Dez cultivares de A. pintoi foram lançados nos últimos 12 anos em diversos países (PAGANELLA, 2002), revitalizando a adoção de leguminosas forrageiras nas regiões tropicais (VALLE, 2001).
A plasticidade no uso de Arachis pintoi como planta forrageira para pastejo direto, para a produção de feno e silagem, para uso paisagístico e em cultivos de cobertura faz desta espécie uma leguminosa ideal para os trópicos latino-americano e mundial (PIZARRO, 2001b). Dez cultivares de A. pintoi foram lançados nos últimos 12 anos em diversos países (PAGANELLA, 2002), revitalizando a adoção de leguminosas forrageiras nas regiões tropicais (VALLE, 2001).
Embora esta espécie seja propagada com eficiência por estolhos, a propagação por sementes facilita a sua mecanização e conseqüente adoção. Entretanto, devido à frutificação geocárpica do gênero Arachis, com 99 % da produção dos frutos ocorrendo nos primeiros 6,0 cm de profundidade do solo (CARVALHO, 1996), a colheita de sementes envolve o revolvimento e peneiramento da camada de solo, elevando o custo de produção (FERGUSON et al., 1992). A maximização da produção de sementes é fundamental para a expansão da área de cultivo.
A variabilidade na produção de sementes do germoplasma de Arachis pintoi é grande, com diferenças que oscilam de zero a sete t/ha (PIZARRO, 2001b). Mesmo entre as cultivares liberadas comercialmente há variações no comportamento reprodutivo. A cv. Belmonte raramente produz sementes, enquanto que as cv. Amarillo, cv. Porvenir e cv. Alqueire são bastante prolíferas (CASTRO et al., 2003). Vários trabalhos buscam explicar estas diferenças. Uma planta que não floresce, não produz sementes, em contraste; o contrário não é garantia de prolificidade (FISCHER, 1999).
No processo de polinização, o pólen maduro é liberado e exposto a um ambiente hostil. É necessário, para que ocorra a germinação do grão de pólen, que este tenha contato com o estigma receptivo (DAFNI, 1992). A diferença entre a produção de sementes de espécies perenes e anuais do gênero Arachis mostrou-se associada à presença de pêlos unicelulares no ápice do estigma, os quais resultam em uma barreira física à polinização (LU et al., 1990). Estes pêlos, com diferenças quanto ao comprimento e densidade, também foram encontrados entre acessos de Arachis pintoi, exclusivamente perenes, mas com atributos diferentes de prolificidade (PEÑALOZA, 1995).
Embora haja indícios de influência da pilosidade da superfície estigmática na produção de sementes, esta associação ainda não foi mensurada. Conhecer as correlações entre os diferentes caracteres envolvidos nas produções de forragem e de sementes, é fundamental para assegurar o êxito em um programa de melhoramento genético (COLLINS, et al., 1997).
O presente trabalho tem como objetivo identificar, em populações F2 de cruzamentos intraespecíficos de Arachis pintoi, as correlações entre os componentes biológicos da produção de forragem e de sementes, com base nos caracteres: comprimento e largura dos folíolos basal e distal, percentagem de cobertura do solo, comprimento do entrenó, espessura e número de estolhos, produção de matéria seca, número de flores/m2 e de sementes/m2, peso médio de sementes, comprimento e densidade de pêlos no ápice do estigma.
Material e métodos
Em dezembro de 2001, mudas de três populações segregantes (Tabela 1) obtidas a partir da colheita de sementes de híbridos intraespecíficos de Arachis pintoi (OLIVEIRA & VALLS, 2002), foram plantadas na Embrapa Cerrados, em Planaltina, DF, localizada a 15o 35’ 30’’ de latitude sul, 47o 42’ 30’’ de longitude oeste e a 1000 m de altitude. O delineamento adotado foi completamente casualizado, com quatro plantas por parcela, e espaçamento de 0,75 m dentro da linha e de 2,0 m entre linhas. Cada planta F2 foi avaliada individualmente.
Tabela 1. Dados de passaporte dos pais envolvidos nos cruzamentos que originaram as populações segregantes F2 de Arachis pintoi avaliadas em campo experimental, em Planaltina, DF. Brasil
| Cruzamento: 13338 x 12787 | ||
| Coletor 1 | Município/Estado/País | Bacia hidrográfica |
| V 13338 | Formosa/GO/Brasil | Paranã |
| GK 12787 | Belmonte/BA/Brasil | Jequitinhonha |
| Cruzamento: 13167 x 12787 | ||
| V 13167 | Unaí/MG/Brasil | São Francisco |
| GK 12787 | Belmonte/BA/Brasil | Jequitinhonha |
| Cruzamento: 13167 x 6791-wf | ||
| V 13167 | Unaí/MG/Brasil | São Francisco |
| V 6791-wf | Formosa/GO/Brasil | Paranã |
1 V = J. F. M. Valls; G = W. C. Gregory; K = A. Krapovickas;
O experimento foi irrigado por aspersão, sendo realizado um corte de uniformização das plantas, 48 dias após o plantio, no dia 21/01/2002. Foram avaliados os seguintes caracteres:
 Cobertura do solo
Cobertura do solo
Medida em percentagem de cobertura por 0,50 m2, seguindo procedimento descrito por TOLEDO (1982), aos 127 dias após o corte de uniformização.
Número de estolhos
Foram contados os números totais de estolhos por planta aos 194 dias após o corte de uniformização.
Produção de matéria seca
Foram realizadas duas avaliações, uma aos 62 dias após o corte de uniformização, no dia 18/03/2002, e outra aos 228 dias após o primeiro cortem, no dia 04/11/2002. O corte foi realizado rente ao solo, removendo toda parte aérea da planta, sendo as amostras pesadas e retiradas sub-amostras para avaliação de matéria seca, sendo estas secas em estufa a uma temperatura de 65 grados centigrados, após 48 horas e pesadas (g).
Número de flores/área: Após o corte de uniformização, semanalmente foi contado o número de flores/área. Para a delimitação da área, foi utilizado um quadrado de dimensões de 0,30 x 0,30 m jogado ao acaso em cima de cada planta.
Pilosidade da superfície estigmática
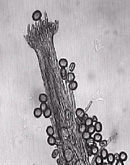 Foram coletadas flores dos genótipos entre 9:00 e 10:00 horas da manhã durante o mês de janeiro de 2002, e fixadas em álcool 70%. Com o auxílio de uma lupa retirou-se o ápice do estigma e montou-se uma lâmina para observar em microscópio óptico com aumento de 20X a densidade e o comprimento dos pêlos da superfície estigmática, sendo os acessos classificados quanto à densidade em poucos (1), intermediários (2) e muitos pêlos (3), e quanto ao comprimento em pêlos longos (3), médios (2) e curtos (1), seguindo metodologia descrita por PEÑALOZA (1995). Foram avaliadas três flores de cada genótipos.
Foram coletadas flores dos genótipos entre 9:00 e 10:00 horas da manhã durante o mês de janeiro de 2002, e fixadas em álcool 70%. Com o auxílio de uma lupa retirou-se o ápice do estigma e montou-se uma lâmina para observar em microscópio óptico com aumento de 20X a densidade e o comprimento dos pêlos da superfície estigmática, sendo os acessos classificados quanto à densidade em poucos (1), intermediários (2) e muitos pêlos (3), e quanto ao comprimento em pêlos longos (3), médios (2) e curtos (1), seguindo metodologia descrita por PEÑALOZA (1995). Foram avaliadas três flores de cada genótipos.
Produção de sementes
Iniciou-se a colheita de sementes no dia 13/11/2002, 300 dias após o corte de uniformização. Para a padronização da área de coleta de sementes foi utilizado um quadrado de 0,50 x 0,50 m, ficando a planta centralizada neste quadrado. Também foi utilizados um outro quadrado de 0,20 x 0,20 m com o objetivo de preservar as partes centrais da planta, diminuindo o risco de perda por morte de acessos. A coleta de sementes foi realizada na área entre o quadrado de 0,50 x 0,50 m e o quadrado de 0,20 x 0,20 m, sendo removido o solo a uma profundidade de 0,15 m e peneirado no campo, repondo o solo no local de onde ele foi retirado, dando condições para as plantas seguirem o seu desenvolvimento. Após, o solo peneirado foi lavada sobre uma peneira e colhidas a semente. Essas foram postas espalhadas em placas e deixadas a temperatura ambiente por cerca de 48 horas, momento em que o excesso de umidade já havia sido eliminado. Os segmentos de frutos foram contadas, separando os segmentos com sementes abortadas ou sem sementes, daqueles com semente, e postos em sacos de papel, sendo pesados e levados para secagem na Embrapa Recursos Genéticos e Biotecnologia em câmara com temperatura de 25oC e umidade entre 7 e 11 %. Semanalmente essas sementes foram pesadas até atingirem peso constante.
Após o corte de uniformização foram marcados com fios de diferentes cores três estolhos por planta, selecionados ao acaso, para as avaliações dos seguintes caracteres:
Espessura do estolho
Foi medida 48 dias após o corte de uniformização, entre o terceiro e o quarto nó, sentido ápice do estolho/base, nos três estolhos marcados por planta, sendo medida com o auxílio de um paquímetro eletrônico, graduado em mm.
Comprimento do entrenó
Foi medido 48 dias após o corte de uniformização, entre o terceiro e o quarto nó, sentido ápice do estolho/base, nos três estolhos marcados por planta, sendo mensurado com um o auxílio de uma régua, graduada em cm.
Tamanho do folíolo
Foi medido, 48 dias após o corte de uniformização, na quarta folha, sentido ápice do estolho/base, dos três estolhos marcados por planta. De cada folha foi medido separadamente o comprimento e a largura do folíolo basal e apical da metade interna da folha, com o auxílio de uma régua, graduada em cm.
Com o objetivo de verificar as associações entre os diferentes caracteres mensurados, foi realizada a analise de correlação simples de Pearson. Foi adotado o procedimento “Proc Corr” do aplicativo computacional de análise de dados SAS (SAS INSTITUTE INC., 1990).
Resultados e discussão
Os resultados mostraram diferenças entre os valores e o nível de significância das correlações nas três populações segregantes. Na maioria das observações, as correlações encontradas na população F2 13338 x 12787, foram, em valor absoluto, superiores às da F2 13167 x 12787 e F2 13167 x 6791-wf, sendo que esta última também apresentou um menor número de correlações estatisticamente significativas. Estas diferenças podem ser em parte explicadas pelas distintas constituições genotípicas de cada população avaliada. Não seria válido calcular um único coeficiente de correlação, para as três populações, uma vez que as variações podem não ser homogênea (STEEL & TORRIE, 1980). Entretanto, é importante salientar que todas as correlações estatisticamente significativas apresentaram, nas três populações segregantes, a mesma orientação, positiva ou negativa.
As correlações com os maiores valores foram encontradas entre o comprimento do folíolo basal e o comprimento do folíolo distal, acima de 0,80 nas três populações. Também foram altas e positivas as correlações envolvendo os caracteres comprimento e largura dos folíolos. Estes caracteres apresentaram correlação positiva com o comprimento do entrenó e a espessura do estolho, acima de 0,6. Da mesma forma, estes dois últimos caracteres apresentaram associação positiva, acima de 0,50, com a percentagem de cobertura do solo.
A alta associação positiva entre o comprimento e largura dos folíolos basal e distal resulta em que, ao selecionar um destes caracteres, o tamanho da folha será alterado. Entre diferentes caracteres vegetativos, em populações de trevo branco, o tamanho do folíolo foi a característica que exerceu maior influência no rendimento de forragem (ANNICCHIARICO, 1993).
Importância também deve ser dada à espessura do estolho, característica muito utilizada em programas de melhoramento genético de trevo branco que visam a aumentar a persistência da pastagem (COLLINS et al, 1997). O número de estolhos, embora também seja um dos maiores indicadores de persistência de uma pastagem em longo prazo (ANNICCHIARICO, 1993; ANNICCHIARICO & PIANO, 1995; PIANO & ANNICCHIARICO, 1995), apresenta a desvantagem, se comparado com a espessura do estolho, de mostrar maior plasticidade fenotípica (COLLINS et al; 1997). Os resultados deste trabalho sugerem que, nas populações avaliadas, também seja importante considerar a espessura do estolho na caracterização e seleção do germoplasma. A associação positiva encontrada entre a espessura do estolho e os componentes biológicos da produção de forragem e o número de sementes/m2, justificam a adoção desse caráter.
As duas avaliações de produção de matéria seca apresentaram correlações positivas entre si, e com os demais caracteres vegetativos avaliados. Com os componentes biológicos da produção de sementes, a primeira avaliação de produção de matéria seca mostrou associação positiva com o número de flores/m2. Correlação negativa foi encontrada entre o segundo corte de avaliação de matéria seca e o peso de sementes, de -0,37 a -0,53, e positiva com o número de sementes/m2, variando de 0,63 a 0,48, sendo que este caráter, também mostrou associação positiva, em torno de 0,40, com os caracteres vegetativos percentagem de cobertura do solo, espessura e comprimento do entrenó.
Os altos valores de produção de matéria seca, percentagem de cobertura do solo e de espessura do estolho, foram compatíveis com os maiores valores de número de sementes/m2, mostrando a possibilidade de se combinar produção de foragem, cobertura do solo e produção de sementes em um único genótipo, o que é bastante desejável. Por outro lado, a associação entre o peso de sementes e o número de sementes foi negativa, - 0,23. É importante mensurar essa associação nas populações de A. pintoi, e buscar enfraquecer tal correlação negativa com a seleção de plantas altamente prolíferas que apresentem maior peso de sementes. Assim, será possível obter sementes maiores, as quais estão relacionado com o maior vigor das plantas nos estágios iniciais de desenvolvimento (HARRIS, 1987).
A ausência de correlação significativa, nas populações avaliadas, entre o número de sementes/m2 e o número de flores/m2, justificam, em parte, situações anteriores onde, embora cortes tenham induzido a fortes picos de florescimento em Arachis pintoi, esta maior produção de flores não resultou em maiores produções de sementes (ARGEL, 1994; ANDRADE & KARIA, 1997). No entanto, dados preliminares sugerem que ao desfolhar drasticamente a pastagem no período de três a seis meses após o plantio, a produção de sementes é bastante reduzida se comparada com cortes feitos após o pico de florescimento, 180 vs 700 kg/ha, respectivamente (PIZARRO, 2001a).
A correlação entre o número de sementes/m2 e o comprimento de pêlos no ápice do estigma foi negativa, -0,45 na F2 13338 x 12787, e -0,63 na F2 13167 x 12787. Porém, nessa última população, a densidade de pêlos na superfície estigmática mostrou associação positiva, 0,25, com o número de sementes/m2. Nas demais populações esta correlação não foi significativa. Esses resultados mostram a influência negativa do comprimento de pêlos no ápice do estigma na produção de sementes, mesmo quando avaliados apenas estigmas do tipo um e dois. Plantas com estigmas do tipo três, tanto para densidade, como para o comprimento, raramente produzem sementes (PEÑALOZA, 1995; OLIVEIRA & VALLS, 2003), não disponibilizando populações segregantes para análise.
A pequena associação positiva entre a densidade de pêlos no ápice do estigma e a produção de sementes mostra que a maior barreira à polinização ocorre devido ao comprimento dos pêlos, e não à quantidade. Esse resultado esta de acordo o tipo de estigma encontrado no cultivar ‘Amarillo', que embora seja classificado com estigma tipo um para o comprimento, e dois para a densidade (PEÑALOZA, 1995), é bastante conhecida por sua alta capacidade de produzir sementes (ARGEL & VALÉRIO, 1993; CRUZ et al., 1999).
Os trabalhos realizados anteriormente ou consideravam a presença ou ausência de pêlos no ápice do estigma como barreira física à polinização, juntamente com o tamanho da superfície estigmática, que acomodaria poucos grãos de pólen (LU et al., 1990). Ou, a associação densidade e comprimento, gerando nove classes de estigma (PEÑALOZA, 1995), mas nunca separando estes dois caracteres e associando-os diretamente com a produção de sementes. Entretanto, é importante salientar que, além do comprimento de pêlos no ápice do estigma, outros mecanismos também podem estar influenciando no sucesso da polinização. Há indícios da presença de auto-incompatibilidade genética nos acessos utilizados como genitores das populações avaliadas neste trabalho (OLIVEIRA & VALLS, 2003). Estudos futuros devem abordar este tema, juntamente com a possibilidade de se vencer a barreira física provocada pela presença de pêlos longos no ápice do estigma. Uma forma possível é através da utilização de insetos polinizadores, como demonstrado em Arachis lignosa (BANKS, 1990).
O presente trabalho elucidou algumas correlações fenotípicas entre os componentes de forragem e de sementes, vindo a ajudar no direcionamento da seleção de características fundamentais em um programa de melhoramento genético de Arachis pintoi, a produção de forragem e de sementes.